Ig重链基因是由V、D、J和C四种不同基因片段所组成。
(一)Ig重链可变区(V区)基因
重链可变区基因是由V、D、J三种基因片段经重排后所形成。
1.重链V区基因的组成 编码重链V区基因长约1000~2000kb,包括V、D、J三组基因片段。
(1)重链V基因片段:小鼠VH基因片段数目为250~1000个。根据VH基因片段核酸序列的相似性(>80%同源性),至少可分为11个家族(family).人V基因片段约为100个,至少可分为6个家族,每个家族含有2~60个成员不等。V基因片段由2个编码区(coding regions)组成:第一个编码区编码大部分信号序列;第二个编码区编码信号序列羧基端侧的4个氨基酸残基和可变区约98个氨基酸残基,包括互补决定区1和2(complementarity determining region 1和2,CDR1和CDR2)。
(2)重链D基因片段:D(diversity)是指多样性。DH基因片段仅存在于重链基因中而不存在于轻链基因。D基因片段编码重链V区大部分CDR3。小鼠DH共有12个片段,位于VH和JH基因片段之间,大部分DH片段较为集中,约占60~80kb,但靠上游的DH可能位于VH区域内,最后一个DH片段与JH基因5'端相距约0.7kb。人类DH片段可能有10~20个左右。
(3)重链的J基因片段:J(joining)指连接,是连接V和C基因片段。JH编码约15~17个氨基酸残基,包括重链V区CDR3除DH编码外的其余部分和第4骨架区。小鼠JH基因片段有4个,与Cμ相距约6.5kb。人有9个JH,其中6个是有功能的JH基因片段。
V、D、J基因片段经重组连接在一起,组成2个外显子,一个外显子编码信号序列的大部分,另一个外显子编码信号序列的其余部分和重链可变区。
小鼠和人在胚系中Ig基因的结构见图3-4和图3-5。
2.重链可变区基因的移位 在重链基因重排开始时,二条染色体上都发生D基因片段移位到J基因片段而发生D-J基因连接。在此以后,只有其中一条染色体上的V基因片段与D-J基因片段连接。VH基因片段5'端含有启动子(promoter),JH和Cμ基因片段之间的内含子中含有转录增强子(transcriptinal enhancer)。如果一条染色体VH基因与D-J基因重排无效(non-productive),另一条染色体的VH基因片段开始发生移位,与D-J基因片段连接。
某些与Ig基因片段重排有关的特殊序列称为识别序列(recognition sequences),位于V基因片段的3'端与J基因片段的5'端之间以及D基因片段的两侧。V基因片段3'端、J基因片段5'端以及D基因片段的两侧也是DNA重排识别信号所在区域,这些识别信号包括三部分:(1)高度保守的回文结构的七聚体(palindromic heptamer);(2)较少保守、富含A/T的九聚体(nonamer);(3)七聚体和九聚体之间不保守的间隔序列(spacer sequence),含有12±1碱基对或23±1碱基对。根据12/23碱基对间隔规则(或称1圈/2圈定律),两个基因片段的重组仅发生在两个基因片段之间:各有一个12个碱基对片段和一个23个碱基对片段的结构(图3-6)。
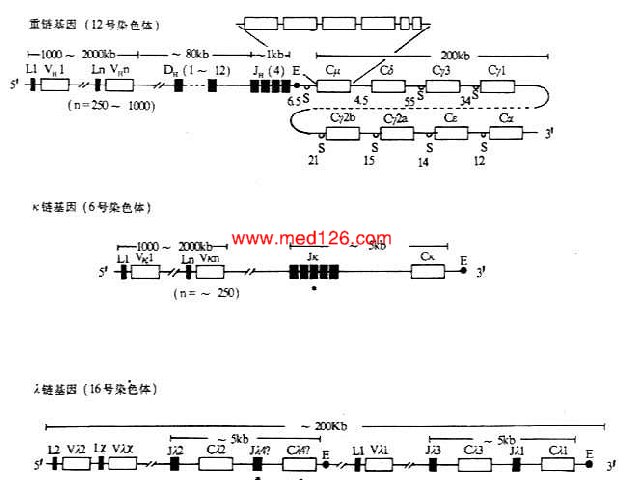
图3-4 小鼠Ig基因结构
注:(1)L:先导序列基因片段 V:可变区基因片段 D:D基因片段
J:J基因片段 C:恒定区基因片段 E:转录增强子
S:转换区 *:假基因
(2)内含子区域所标数字表示DNA长度(kb)。
(3)每个CH基因用一个方框表示,实际上包括几个外显子,如Cμ含有6个外显子。
(4)λ基因增强子位于Cλ4和Cλ1基因片段的下游。
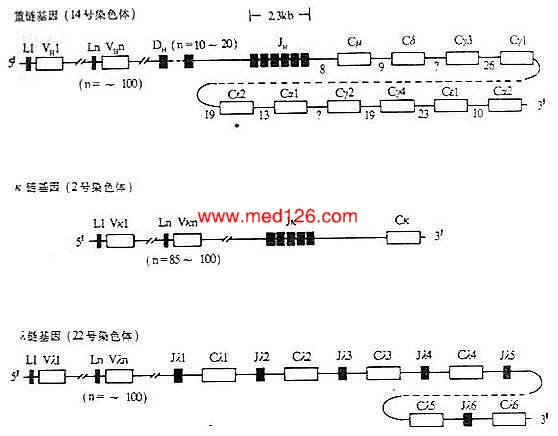
图3-5 人Ig基因结构
注:(1)L:先导序列基因片段 V:可变区基因片段 D:D基因片段
J:J基因片段 C:恒定区基因片 *:假基因
(2)内含子区域所标数字表示DNA长度(kb)
(3)每个CH基因用一个方框表示,实际上包括几个外显子
参与V/(D)/J基因重组过程的酶称为V/(D)/J重组酶(recombinase),有关于执行识别、切割和重新连接基因片段重组酶的纯化和鉴定工作还刚开始。重组酶实际上包括重组过程中多种酶的活性。最近在前B细胞(pre-b cell)中已经鉴定出两种刺激Ig基因重排的基因,称为重组激活基因1(recombination activating gene 1,RAG-1)和重组激活基因2(RAG-2),其确切的作用机理还不太清楚。重组酶作用的特点是:(1)淋巴细胞特异性的,非淋巴样细胞如成纤维细胞无重组酶活性,这可能解释了Ig基因的重排仅见于B淋巴细胞。目前一般认为T细胞TCR基因重排中的重组酶与B细胞中重组酶相同或相似。(2)重组酶发挥其功能仅限于B细胞发育早期,未成熟B细胞如前B细胞(pre-b cell)细胞系重组酶活性很高,但抗体生成细胞或骨髓瘤细胞无明显重组酶活性,因此时B细胞已经分泌某一特异性抗体,不再发生重排其它的Ig基因,因此也不会改变原先所产生抗体的特异性。转换重组酶(switch recom-binase)可能与VDJ重组酶(VDJ recombinase)相似,但缺乏七聚体/九聚体(heptamer/nonamer)识别蛋白。重组酶功能异常可导致机体不能产生Ig和TCR,很可能与重症联合免疫缺陷(severe combined immunodeficiency,SCID)的发生有关。例如SCID小鼠16号染色体着丝点末端存在一个scid基因,为单基因常染色体遗传基因。scid基因纯合将影响DNA重组酶的识别功能,在TCR或BCR基因片段重排时不能识别正确的位点,使T细胞、B细胞在淋巴干细胞发育早期即夭折,导致重症联合免疫缺陷。

图3-6 参与Ig基因重排组酶识别的DNA序列




