二、由膜的特异受体蛋白质、G-蛋白和膜的效应器酶组成的跨膜信号传递系统
这是另一类型的跨膜信号传递。最初是从对激素作用机制的研究开始的。60年代在研究肾上腺素引起肝细胞中糖原分解为葡萄糖的作用机制时,发现如果使肾上腺素单独和分离出的细胞膜碎片相互作用,可以生成一种分子量小、能耐热的物质,当把这种物质同肝细胞的胞浆单独作用时,也能引起其中糖原的分解,同肾上腺素作用于完整的肝细胞时有类似的效应。实验提示,在肾上腺素正常起作用时,它只是作用于肝细胞的膜表面。通过某种发生在膜结构中的过程,先在胞浆中生成一种小分子物质,后者再实现肾上腺素分解糖原的作用。这种小分子物质不久被证明是环-磷酸腺苷(即cAMP,环磷腺苷)。以后又陆续发现,很多其他激素类物质作用于相应的靶细胞时,都是先同膜表面的特异受体相结合,再引起膜内侧胞浆中cAMP含量的增加(有时是它的减少),实现激素对细胞内功能的影响。这样就把cAMP称作第二信使,这是相对于把激素分子这类外来化学信号看作第一信使而言的。
导致cAMP产生的膜结构内部的过程颇为复杂:它至少与膜中三类特殊的蛋白质有关。第一类是能与到达膜表面的外来化学信号作特异性结合的受体蛋白质,这是一些真正可以称作受体的物质。目前已用分子生物学的方法证明,它们是一些独立的蛋白质分子;已经确定的近100种这类受体,都具有类似的分子结构,也属于同一蛋白质家族:即它们都由约300~400个氨基酸残基组成,有一个较长的细胞外N-末端,接着在肽链中出现7个由22~28个主要为疏水性氨基酸组成的α-螺旋,说明这肽链至少要反复贯穿膜7次,形成一个球形蛋白质分子,还有一段位于膜内侧的肽链C-末端。目前认为,受体分子中第7个跨膜螺旋是能够识别、即能结合某种特定外来化学信号的部位;在受体因结合了特异化学信号而激活时,将进而作用于膜中另一类蛋白质,即G-蛋白质。
G-蛋白是鸟苷酸结合蛋白(guanine nucleotide-binding protein)的简称,也是存在于膜结构中的一类蛋白质家族,根据它们分子结构中少数氨基酸残基序列上的不同,已被区分出有数十种,但结构和功能极为相似。G-蛋白通常由α-、β-、和γ-3个亚单位组成;α-亚单位通常起催化亚单位的作用,当G-蛋白未被激活时,它结合了一分子的GDP(二磷酸鸟苷);当G-蛋白与激活了的受体蛋白在膜中相遇时,α-亚单位与GDP分离而又与一分子的GTP(三磷酸鸟苷)结合,这时α-亚单位同其他两个亚单位分离,并对膜结构中(位置靠近膜的内侧面)的第三类称为膜的效应器酶的蛋白质起作用,后者的激活(或被抑制)可以引致胞浆中第二信使物质的生成增加(或减少)。上述肾上腺素的作用,就是先由激素激活膜上相应的受体后,通过一种称为Gs(兴奋性G-蛋白)的G-蛋白的中介,激活了作为效应器酶的腺苷酸环化酶(图2-9箭头1),使胞浆中的ATP生成了起第二信使作用的cAMP(图2-9中箭头2)。由于第二信使物质的生成经过多级催化作用,少数几个膜外化学信号分子同受体的结合,就可能在胞浆中生成数目众多的第二信使分子,这是这种类型的跨膜信号传递的重要特点之一。
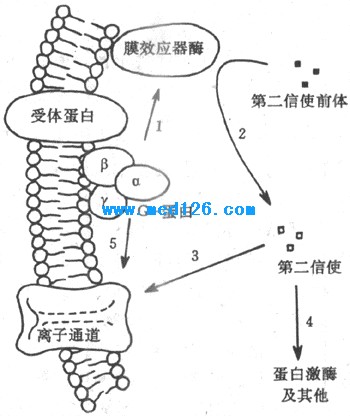
图2-9 由膜受体-G-蛋白-膜效应器酶组成的跨膜信号
传递系统和第二信使类物质的生成
目前发现膜的效应器酶并不只腺苷酸环化酶一种,因而第二信使物质也不只cAMP一种,如近年来还发现,有相当数量的外界刺激信号作用于受体后,可以通过一种称为Go的G-蛋白,再激活一种称为磷脂酶C的膜效应器酶,以膜结构中称为磷脂酰肌醇的磷脂分子为间接底物,生成两种分别称为三磷酸酰肌醇(IP3)和二酰甘油(DG)的第二信使,影响细胞内过程,完成跨膜信号传递。虽然如此,对应于细胞所能接受的多种刺激和与它们相对应的受体数目而言,膜内G-蛋白、效应器酶和最后生成的第二信使类物质的种类,还是相对地少得多。这说明,上述由膜中蛋白质酶促反应生成第二信使的途径,具有相当程度的“通用”性质。
由于上述这种跨膜信号传递的形式是在研究激素的作用机制时发现的,而且后来发现绝大多数肽类激素都是通过这一形式起作用的,因此曾一度错误地认为,这只是激素性化学信号跨膜信号传递方式。但近年的资料说明,事实并非如此:在神经递质类物质中,除了上述氨基酸类递质外,其余不论是小分子的经典递质还是后来发现的数量众多的神经肽类物质(目前已近50种),都主要是以在突触后细胞中产生第二信使类物质来完成跨膜信号传递的,这些第二信使物质通过在胞浆中的扩散,在膜的内侧面作用于某些特殊的离子通道(图2-9中箭头3),引起突触后膜较广泛而缓慢的电变化。最近证明,在视网膜信号转换过程中,光量子被作为受体的视色素如视紫红质(也具有7个跨膜α-螺旋的结构特点)吸收后,也是先激活称为Gt(转换蛋白)的G-蛋白,再激活作为效应器的磷酸二酯酶,使视杆细胞外段中cGMP的分解加强,最后使光刺激转变为外段膜的电变化(见第九章)。




