6.未分化的间充质细胞 未分化的间充质细胞(undifferentiated mesenchymal cell)是保留在成体结缔组织内的一些较原始的细胞,它们保持着间充质细胞的分化潜能,在炎症与创伤时可增殖分化为成纤维细胞、脂肪细胞。间充质细胞常分布在小血管尤其是毛细血管周围,并能分化为血管壁的平滑肌和内皮细胞。
7.白细胞血液内的白细胞,受趋化因子的吸引,常穿出毛细血管和微静脉,游走到疏松结缔组织内,行使其功能,参与免疫应答和炎症反应。疏松结缔组织内以嗜酸性粒细胞、淋巴细胞、中性粒细胞多见。游走出的单核细胞将分化为巨噬细胞。
(二)纤维
1.胶原纤维 胶原纤维(collagenous fiber)数量最多,新鲜时呈白色,有光泽,又名白纤维。He 染色切片中呈嗜酸性,着浅红色。纤维粗细不等,直径1-20μm,呈波浪形,并互相交织。胶原原纤维由直径20~200nm的胶原原纤维粘合而成(图3-2)。电镜下,胶原原纤维显明暗交替的周期性横纹,横纹周期约64nm(图3-12)。胶原纤维的韧性大,抗拉力强。胶原纤维的化学成分为Ⅰ型和Ⅱ型胶原蛋白。胶原蛋白(简称胶原,collagen)主要由成纤维细胞分泌。分泌到细胞外的胶原再聚合成胶原原纤维,进而集合成胶原纤维。
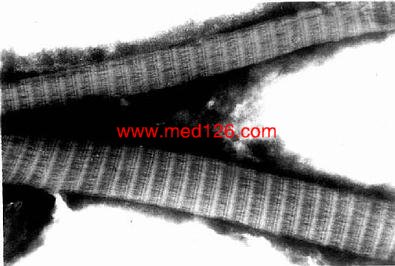
图3-12 人胶原纤维电镜像×150000
(同济医科大学武忠弼教授供图)

图3-13胶原纤维与基质形成过程示意图
胶原纤维形成的基本过程如下(图3-13):
(1)细胞内合成前胶原蛋白分子:成纤维细胞摄取合成蛋白质所需的氨基酸,包括脯氨酸、赖氨酸和甘氨酸,在粗面内质网的核糖体上按照特定的胶原mRNA的碱基序列,合成前α-多肽链。后者边合成边进入粗面内质网腔内,并在羟化酶的作用下,将肽链中的脯氨酸和赖氨酸羟化。经羟化后,三条前α-多肽链互相缠绕成绳索状的前胶原蛋白分子(procollagen molecule)。溶解状态的前胶原蛋白分子,两端未缠绕,呈球状构型,在粗面内质网腔内或转移到高尔基复合体内加入糖基后,分泌到细胞外。
(2)原胶原蛋白分子的细胞外聚合:细胞外的前胶原蛋白分子,在肽内切酶的作用下,切去分子两端球状构形部分,形成原胶原蛋白分子(tropocol-lagen)粗约1.5nm,长约300nm。原胶原蛋白分子平行排列聚合成胶原原纤维。聚合时,相互平行的相邻分子错开1/4分子长度,同一排的分子,首尾相对并保持一定距离,聚合成束,于是形成具有64nm周期横纹的胶原原纤维。聚合时,分子内、分子间的化学基因进行缩合、交联,增加原纤维的稳固性。若干胶原原纤维经糖蛋白粘合成粗细不等的胶原纤维。
胶原纤维的一菜成受多方面的影响和调控。如细胞内脯氨酸的含量直接影响前α-多肽链的合成。缺氧或缺乏维生素C或Fe2+等辅助因子,导致前α-多肽链的羟化受到抑制,造成前胶原蛋白合成障碍,影响创伤的愈合。聚合时,如胶原蛋白分子内和分子间的交联障碍(常因赖氨酰氧化酶不足所致)将影响胶原纤维的稳固性。除成纤维细胞外,成骨细胞、软骨细胞、某些平滑肌细胞等起源于间充质的细胞以及多种上皮细胞也能产生胶原蛋白。
不同组织的胶原蛋白其分子类型不同,已证实α-多肽链按其一级结构分为α1,α2,α3,三类,各类又分为10型,如α1(Ⅰ)、α1(Ⅱ)、α1(Ⅲ)、α1(Ⅲ)……α1(X)。
根据构成胶原蛋白三股肽链的不同,现已发现有11种不同类型的胶原。现将主要几种类型的组成、分布和特点列举于表(表3-1)。
表3-1 胶原蛋白的类型、分布和特点
| 类型 | 前胶原蛋白的三股肽链 | 分 布 | 主 要 特 点 |
| Ⅰ | [α1(Ⅰ)]2α2(Ⅰ) | 真皮、筋膜、巩膜、被膜、腱、纤维软骨、骨、牙本质 | 构成致密并有横纹的粗纤维束,抗拉力强 |
| Ⅱ | [α1(Ⅱ)]3 | 透明软骨和弹性软骨 | 构成有横纹的细原纤维,抗压力较强 |
| Ⅲ | [α1(Ⅲ)]3
[α1(Ⅳ)]2α2(Ⅳ) |
网状纤维、平滑肌、神经内膜、动脉、肝、脾、肾、肺、子宫 | 构成有横纹的细原纤维,维持器官的形态结构 |
| Ⅳ | [α1(Ⅳ)]3
[α2(Ⅳ)]3 [α1(Ⅴ)]2α2(Ⅴ) |
基膜基板、晶 状体囊 | 不形成原纤维,为均质状膜,支持和滤过作用 |
| Ⅴ | [α1(Ⅴ)]3
α1(Ⅴ)α2(Ⅴ)α3(Ⅴ) |
胎膜、肌、腱鞘 | 构成细的无横纹原纤维 |
2.弹性纤维 弹性纤维(elastic fiber)新鲜状态下呈黄色,又名黄纤维。在HE标本中,着色轻微,不易与胶原纤维区分。但醛复红(aldehyde-fuchsin)或地衣红(orcein)能将弹性纤维染成紫色或棕褐色。弹性纤维较细,直行,分支交织,粗细不等(0.2-1.0μm),表面光滑,断端常卷曲(图3-2)。电镜下,弹性纤维的核心部分电子密度低,由均质的弹性蛋白(elastin)组成,核心外周覆盖微原纤维(microfibril),直径约10nm。弹性蛋白分子能任意卷曲,分子间藉共价键交联成网。在外力牵拉下,卷曲的弹性蛋白分子伸展拉长;除去外力后,弹性蛋白分子又回复为卷曲状态(图3-14)。
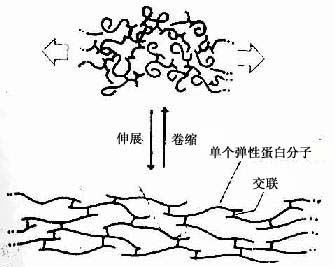
图3-14 伸缩状态下弹性蛋白分子的构形变化
弹性纤维富于弹性而韧性差,与胶原纤维交织在一起,使疏松结缔组织既有弹性又有韧性,有利于器官和组织保持形态位置的相对恒定,又具有一定的可变性。
3.网状纤维 网状纤维(reticular fiber)较细,分支多,交织成网。网状纤维由Ⅲ型胶原蛋白构成,也具有64nm周期性横纹。纤维表面被覆蛋白多糖和糖蛋白,故PAS反应阳性,并具嗜银性。用银染法,网状纤维呈黑色,故又称嗜银纤维(argyrophil fiber)。网状纤维多分布在结缔组织与其它组织交界处,如基膜的网板、肾小管周围、毛细血管周围。在造血器官和内分泌腺,有较多的网状纤维,构成它们的支架。
(三)基质
基质(ground substance)是一种由生物大分子构成的胶状物质,具有一定粘性。构成基质的大分子物质包括蛋白多糖和糖蛋白。
蛋白多糖(proteoglycan)是由蛋白质与大量多糖结合成的大分子复合物,是基质的主要成分。其中多糖主要是透明质酸(hyaluronic acid),其次是硫酸软骨素A 、C(chondroitin sulfate A、C)、硫酸角质素A、C(keratin sulfate)硫酸乙酰肝素(heparan sulfate)等。它们都是以含有氨基已糖的双糖为基本单位聚合成的长链化合物,总称为糖胺多糖(glycosaminoglycan,GAG)。由于糖胺多糖分子存在大量阴离子,故能结合大量水(结合水)。透明质酸是一种曲折盘绕的长链大分子,拉直可达2.5μm,由它构成蛋白多糖复合物的主干,其它糖胺多糖则以蛋白质为核心构成蛋白多糖亚单位,后者再通过连接蛋白结合在透明质酸长链分子上(图3-15)。蛋白多糖复合物的立体构型形成有许多微孔隙的分子筛,小于孔隙的水和溶于水的营养物、代谢产物、激素、气体分子等可以通过,便于血液与细胞之间进行物质交换。大于孔隙的大分子物质,如细菌等不能通过,使基质成为限制细菌扩散的防御屏障。溶血性链球菌和癌细胞等能产生透明质酸酶,破坏基质的防御屏障,致使感染和肿瘤浸润扩散。

图3-15 蛋白多糖分子结构模型
糖蛋白(glycoprotein)是基质内另一类重要的生物大分子,与蛋白多糖相反,其主要成分是蛋白质。从基质内已经分离出多种糖蛋白,主要的有纤维粘连蛋白(fibronectin FN)层粘连蛋白(laminin)和软骨粘连蛋白(chondronectin)等。这类基质大分子不仅参与基质分子筛的构成,同时通过它们的连接和介导作用也影响细胞的附着和移动以及参与调节细胞的生长和分化。
纤维粘连蛋白是基质中一种重要的糖蛋白,存在于胶原纤维和许多结缔组织细胞周围。在电镜下,纤维粘连蛋白呈原纤维状,由两条多肽链组成,两条肽链的一端由若干二硫键连接。每一肽链上均有若干特定的功能区,能分别与细胞、胶原、肝素和纤维素等结合。于是,纤维粘连蛋白作为一种中介蛋白,能将细胞连接到胶原、肝素等细胞外基质上。
组织液(tissue fluid)是从毛细血管动脉端渗入基质内的液体,经毛细血管静脉端和毛细淋巴管回流入血液或淋巴,组织液不断更新,有利于血液与细胞进行物质交换,成为组织和细胞赖以生存的内环境。当组织液的渗出、回流或机体水盐、蛋白质代谢发生障碍时,基质中的组织液含量可增多或减少,导致组织水肿或脱水。




